Tannhjul i biologi, bevis på design?!
Av Otangelo Grasso, hentet via FB -lenke.
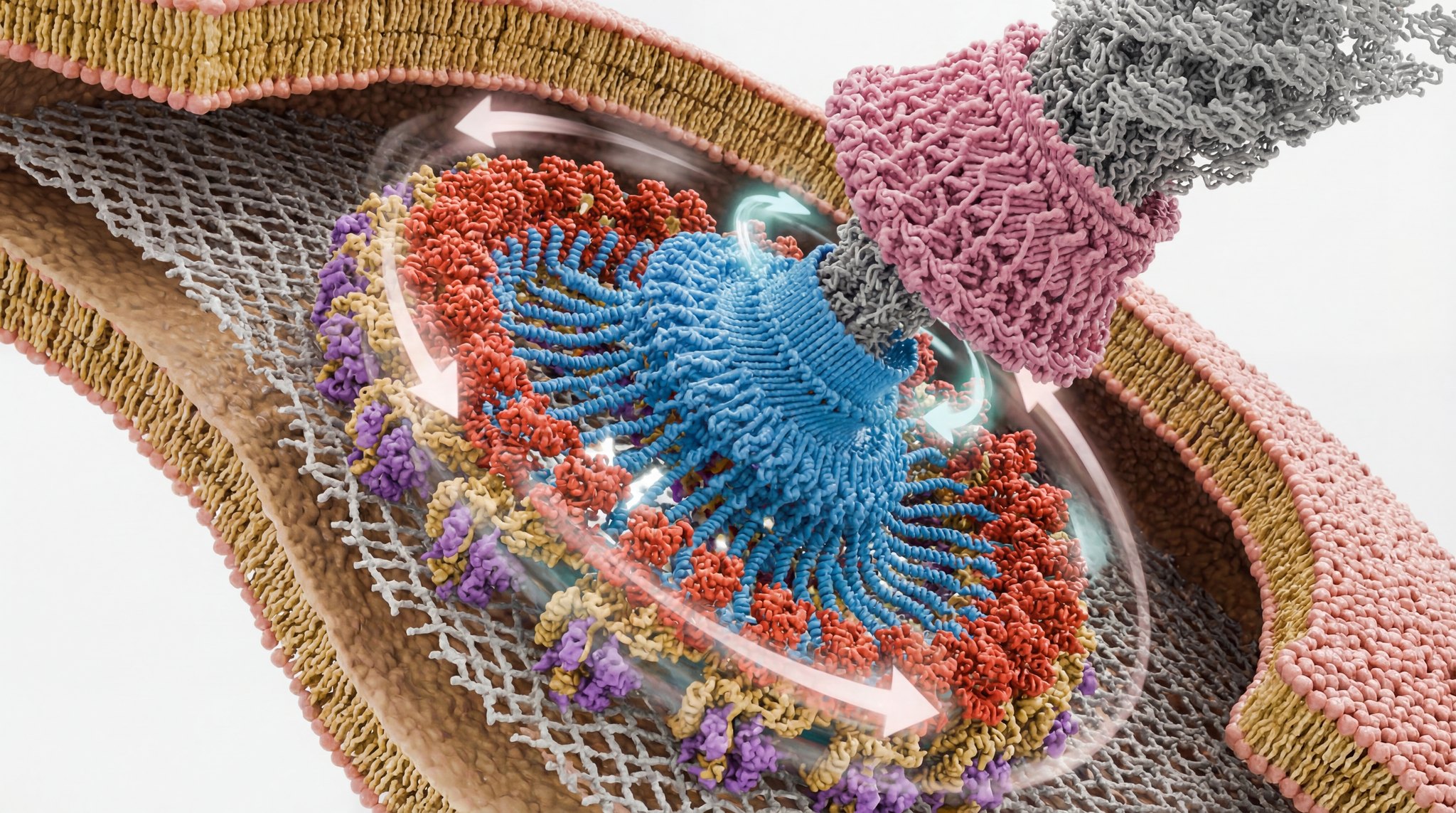 Momentgenererende enheter i den bakterielle flagellmotoren er rotasjonsmotorer (Hosu og to andre Harvard-fysikere, PNAS, publisert 12/10/25 -lenke).
Momentgenererende enheter i den bakterielle flagellmotoren er rotasjonsmotorer (Hosu og to andre Harvard-fysikere, PNAS, publisert 12/10/25 -lenke).
Bilde 1. Gir med tannhjul i levende orgnaisme (Kredit O. Grasso.)
Betydning
Bakterier svømmer ved å rotere stive spiralformede flagellfilamenter. Her finner vi at den momentgenererende enheten som driver flagellrotasjonen i seg selv er en rotasjonsmotor. Hver momentgenererende enhet er en heteromer makromolekylær maskin - en pentamer av MotA-underenheter som omgir en dimer (smsatt. fra to enheter) av protonledende MotB-underenheter. Dreiemoment genereres når MotA spinner rundt MotB. MotA-pentameren samhandler med rotoren til flagellmotoren på en måte som ligner sammenflettede tannhjul. Den bakterielle flagellmotoren drives av det første settet med innviklede tannhjul som er beskrevet i noen levende celle.
Det denne nye artikkelen gjør er å stramme inn analogien mellom flagellmotoren og en designet roterende maskin til den nesten brister.
Hosu og kolleger viser at hver statorenhet ( ) ikke bare er et statisk dreiemoment-transduserende stillas, men en roterende motor i seg selv. Ved å bruke polarisert fotobleking på motorer som kjører in vivo, sporer de merkede MotA og MotB og finner ut at MotB-dimeren forblir låst til celleveggen mens MotA-pentameren spinner rundt den, og roterer omtrent 6,2 ganger raskere enn hovedrotoren. Det er akkurat det du forventer av et tannhjul: et lite tannhjul som driver et større med et definert girforhold.
) ikke bare er et statisk dreiemoment-transduserende stillas, men en roterende motor i seg selv. Ved å bruke polarisert fotobleking på motorer som kjører in vivo, sporer de merkede MotA og MotB og finner ut at MotB-dimeren forblir låst til celleveggen mens MotA-pentameren spinner rundt den, og roterer omtrent 6,2 ganger raskere enn hovedrotoren. Det er akkurat det du forventer av et tannhjul: et lite tannhjul som driver et større med et definert girforhold.
 Dette er bygget på strukturelt arbeid som viser at statoren har en asymmetrisk 5:2-arkitektur, en MotB-"aksel" omgitt av et MotA-"hjul". En slik konfigurasjon hadde allerede antydet mulig rotasjon av statoren; denne artikkelen måler den direkte og kvantifiserer forholdet mellom stator- og rotorhastigheter.
Dette er bygget på strukturelt arbeid som viser at statoren har en asymmetrisk 5:2-arkitektur, en MotB-"aksel" omgitt av et MotA-"hjul". En slik konfigurasjon hadde allerede antydet mulig rotasjon av statoren; denne artikkelen måler den direkte og kvantifiserer forholdet mellom stator- og rotorhastigheter.
Så vi har nå, inne i en autonom celle, et protondrevet rotasjonssystem der:
*en MotB-dimer forankres i peptidoglykanen (peptid: aminosyrer; glykan-sukkerkjeder -hovedkomponent i bakteriers cellevegg);
*en MotA-pentamer danner en roterende nanoring rundt den;
*protonstrøm gjennom spesifikke ionebindende rester driver trinnvis rotasjon av denne ringen;
*den ringen går i inngrep med C-ringen til den flagellære rotoren "som sammenflettede tannhjul", og overfører dreiemoment for å spinne det lange spiralformede filamentet.
For en forkjemper for intelligent design er flere trekk her slående.
Bilde 2. Gir inni gresshoppe
For det første er arkitekturen funksjonelt integrert. 5:2-statorgeometrien, formen og avstanden til MotA-tennene som griper inn i FliG på rotoren, protonbindingskjemien i MotA og MotB, og den mekaniske stivheten til MotB-ankeret må alle justeres sammen slik at protonfluksen omdannes til et jevnt roterende MotA-hjul, som deretter driver en større rotor med et fast girforhold. Hvis protonkanalen brytes, kontaktflatene endres, eller den stive forankringen mistes, slutter systemet å fungere som en roterende momentgenerator. Delene er arrangert i samarbeidende systemer snarere enn løst koblede elementer.
For det andre er analogien til konstruerte enheter ikke overfladisk. I flere tiår har lærebøker allerede beskrevet den bakterielle flagellarmotoren som en "rotasjonsmotor" eller "proteinturbin", drevet av en ionegradient over membranen. Det denne artikkelen legger til er bokstavelige tannhjul: et lite, raskt roterende MotA-hjul som driver en større, langsommere C-ring via mekanisk sammenkobling. Innen menneskelig teknologi, når vi ser et flerdelt system med stator, rotor, spesifikt girforhold og energitilførsel gjennom en ion/elektrisk gradient, klassifiserer vi det uten å nøle som en konstruert motor. Her ser vi et sammenlignbart skjema utført på nanometerskala av bittesmå, intrikat konstruerte molekylære maskiner. Korrespondansen mellom biologiske og menneskeskapte rotasjonssystemer blir mye mer detaljert.
 For det tredje er statorens drift ikke bare mekanisk, den styres av kraftige molekylære kontrollnettverk. MotAB-montering i motoren avhenger av membranpotensial; statorenheter rekrutteres og frigjøres dynamisk avhengig av belastning; mutasjoner i spesifikke ladede rester kan avskaffe dreiemomentgenerering mens andre aspekter av proteinet forblir intakte. Det betyr at cellens reguleringssystem må "vite" når nok MotAB-girenheter er installert, modulere antallet deres og koordinere deres funksjon med resten av livets molekylære arbeidsstyrke, proteomet og membranarkitekturen som opprettholder protonmotorkraft og integrerer kjemotaksisignaler.
For det tredje er statorens drift ikke bare mekanisk, den styres av kraftige molekylære kontrollnettverk. MotAB-montering i motoren avhenger av membranpotensial; statorenheter rekrutteres og frigjøres dynamisk avhengig av belastning; mutasjoner i spesifikke ladede rester kan avskaffe dreiemomentgenerering mens andre aspekter av proteinet forblir intakte. Det betyr at cellens reguleringssystem må "vite" når nok MotAB-girenheter er installert, modulere antallet deres og koordinere deres funksjon med resten av livets molekylære arbeidsstyrke, proteomet og membranarkitekturen som opprettholder protonmotorkraft og integrerer kjemotaksisignaler.
Fra et designperspektiv ser dette ut som et ingeniørmessig vidunder av høyteknologi bygget av proteiner: en nanoskala girkasse drevet av transmembrane ionestrømmer, boltet (konseptuelt, ikke bokstavelig) inn i celleveggen, spunnet av cellens elektrokjemiske "batteri", og koblet til en lang spiralformet propell for å generere skyvekraft. Den kjemiske arkitekturen til MotA og MotB legemliggjør et komplekst spesifisert mønster: spesifikke aminosyrer på presise posisjoner danner protonkanaler, hengselområder og tannlignende grensesnitt som sammen implementerer en målrettet prosess, og omdanner protonstrøm til kontrollert rotasjon.
Bilde 3. Leibniz så det
Hvor kommer intelligent designresonnement inn? Den bruker et veldig enkelt slutningsmønster:
1. I vår uniforme og gjentatte erfaring oppstår rotasjonsmotorer med statorer, rotorer og tannhjul fra sinn, ikke fra ikke-styrte fysiske prosesser.
2. Den bakterielle flagellære motoren viser seg nå å ikke bare være "motorlignende", men å ha interne rotasjonsstatorer som går i inngrep med  rotoren, lik faktiske tannhjul, med et definert girforhold.
rotoren, lik faktiske tannhjul, med et definert girforhold.
3. Derfor er det, ved analogi og ved å appellere til kjente årsaker, rimelig å behandle dette systemet som en kandidat for artefakthypotesen: at informasjonen som spesifiserer det i genomet stammer fra et intellekt, ikke fra ren ikke-biologisk tilfeldighet og nødvendighet.
Dette betyr ikke at PNAS-artikkelen i seg selv argumenterer for design; det gjør den ikke. Forfatterne arbeider fullt ut innenfor et naturalistisk rammeverk og fokuserer på mekanisme, ikke opprinnelse. Det betyr heller ikke at neodarwinistiske forklaringer er logisk umulige. Evolusjonsbiologer vil fortsette å foreslå trinnvise scenarier, genduplikasjoner og koopsjon fra enklere ionekanaler eller sekresjonssystemer for å bygge opp den moderne motoren. Disse forslagene må imidlertid ta hensyn til en girbasert dreiemoment-mekanisme med spesifikk symmetri (5:2), et kvantifisert girforhold, protonkoblet trinnkjemi, monteringssjekkpunkter og integrering i det større cellulære kontrollnettverket.
Bilde 4. Om aliens finnes, vil de kunne ha mye å si om mennesker og maskiner
Så opprinnelsesspørsmålet blir nå skarpere, ikke mer uklart. Man må forklare ikke bare "en spinnende ting", men en stator som selv spinner rundt en fast aksel, engasjerer en rotor lik sammenflettede tannhjul, og gjør dette inne i en membran samtidig som den opprettholder protongradienter og synkroniserer med andre motorer på samme celle. Hver av disse begrensningene innsnevrer rommet for levedyktige baner. Sannsynligheten for å snuble over et slikt system ved ustyrt tukling i sekvensrommet blir mer skremmende usannsynlig etter hvert som de tekniske detaljene multipliseres.
 Bilde 5. Om maskin-analigoen isolerees..
Bilde 5. Om maskin-analigoen isolerees..
For et intelligent designperspektiv fungerer dette nye resultatet som bekreftende bevis. Det forsterker synspunktet om at vi ikke ser på en vag, generisk virvel av molekyler, men på nøye utformede løsninger som implementerer en svært spesifikk mekanisk logikk. ATGC-kvartetten i DNA-et bærer instruksjonene for å bygge MotA, MotB, rotorproteinene, eksportapparatet, foringer, lagrene og kjemotaksinettverket, alt arrangert i en funksjonelt integrert helhet. Informasjon benyttes til å organisere alt til en sammenhengende nanomaskin hvis virkemåte ligner teknologier som vi i alle andre sammenhenger tilskriver sinn.
Metodisk sett har vitenskapen her gjort jobben sin: den har avdekket mer utsøkte designdetaljer ned til nesten atomær oppløsning. Intelligent design spør deretter om disse høypresisjons-nanogirkassene best kan forklares som tilfeldige biprodukter av blinde fysiske prosesser, eller som planlagte oppfinnelser av en kognitiv agent. Hosu et al.-artikkelen avgjør ikke det filosofiske spørsmålet, men den beriker absolutt det faktagrunnlaget som debatten nå må fortsette på.
Referanser:
Momentgenererende enheter i den bakterielle flagellmotoren er rotasjonsmotorer
Frontiers in Microbiology Artikkel
Den roterende bakterielle flagellmotoren
Motilitetsproteininteraksjoner i den bakterielle flagellmotoren
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund